











以下文章來源于BioArt ,作者Qi

BioArt
高屋建瓴,提供專家點(diǎn)評(píng),引導(dǎo)學(xué)術(shù)爭(zhēng)論,展現(xiàn)學(xué)術(shù)批評(píng);誠(chéng)心實(shí)意,關(guān)注科研生態(tài),推廣科研經(jīng)驗(yàn),倡導(dǎo)師生交流。

關(guān)注我們,獲取更多相關(guān)資訊
結(jié)直腸癌(Colorectal cancer, CRC)是全世界范圍內(nèi)與癌癥相關(guān)死亡的第二大主要原因,尤其是在50歲以下的個(gè)體中其患病率正逐步攀升。CRC作為腸道上皮的異質(zhì)性疾病,其特征在于突變的累積和免疫反應(yīng)的失調(diào)。越來越多的證據(jù)表明,高達(dá)90%的疾病風(fēng)險(xiǎn)是由飲食等環(huán)境因素所致,這與近些年來提出CRC相關(guān)“致癌菌群”的文獻(xiàn)報(bào)道相一致。然而,與眾所周知的幽門螺桿菌在胃癌中的因果作用不同,目前尚未鑒定觸發(fā)CRC的特定微生物。這種腸道菌群失調(diào)是導(dǎo)致CRC的直接原因還是僅僅代表“旁觀者效應(yīng)(bystander effect)”尚不清楚。因此,破譯驅(qū)動(dòng)CRC的特定致病菌群或代謝產(chǎn)物并表征其潛在機(jī)制十分必要。
近日,來自牛津大學(xué)的Fiona Powrie課題組在Nature 雜志上發(fā)表了一篇題為“Host–microbiota maladaptation in colorectal cancer” 的綜述,系統(tǒng)回顧了截至目前已報(bào)道的CRC中宿主與腸道菌群的相互作用,重點(diǎn)分析與腸道屏障破壞,基因毒性和炎癥反應(yīng)的有關(guān)機(jī)制,這一領(lǐng)域存在的未知和挑戰(zhàn)以及新興技術(shù)的應(yīng)對(duì)策略。
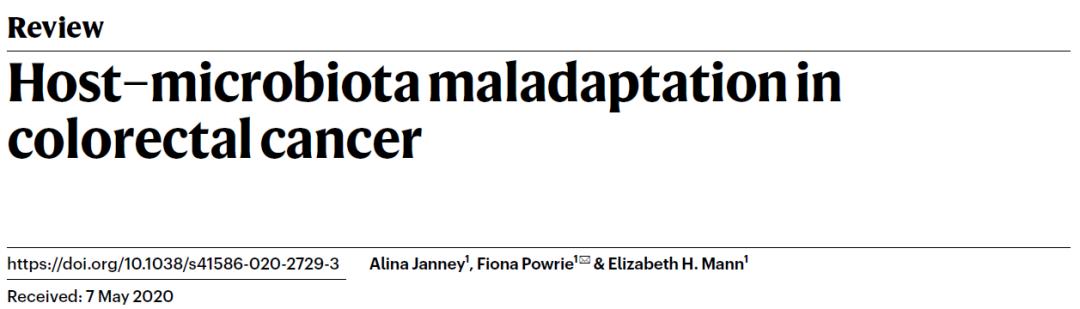
一、宿主建立對(duì)腸道菌群的防御屏障
長(zhǎng)期以來,哺乳動(dòng)物結(jié)腸與多樣化的微生物生態(tài)系統(tǒng)共同進(jìn)化,在一定程度上通過在宿主中形成強(qiáng)大的免疫系統(tǒng),以促進(jìn)宿主和微生物群落的互利共贏。維持這種平衡的關(guān)鍵是腸道上皮細(xì)胞(intestinal epithelial cells, IEC),它們?cè)谒拗?菌群交界面上形成單層物理屏障,并由位于腸隱窩(intestinal crypts)中的多能性腸干細(xì)胞(multipotent intestinal stem cells)不斷給予補(bǔ)充。IEC的增殖受干細(xì)胞龕(stem cell niche)的調(diào)節(jié),并涉及底層間充質(zhì)的旁分泌信號(hào)傳導(dǎo),而上皮間緊密連接蛋白與保護(hù)性粘蛋白的分泌協(xié)同增強(qiáng)屏障的完整性。
除去物理屏障的作用,IEC還通過模式識(shí)別受體來感知微生物群。這些受體的刺激促進(jìn)上皮細(xì)胞修復(fù)和緊密連接蛋白的上調(diào),并觸發(fā)將信號(hào)傳遞至固有層細(xì)胞的多種細(xì)胞因子產(chǎn)生。至關(guān)重要的是,上皮細(xì)胞并非獨(dú)立存在,而是與免疫細(xì)胞和間充質(zhì)一起形成“三元組(triad)”。如果微生物或其產(chǎn)物破壞了上皮屏障防線,那么免疫細(xì)胞和間充質(zhì)細(xì)胞將通過啟動(dòng)一系列支持上皮完整性的信號(hào)網(wǎng)絡(luò)來充當(dāng)?shù)诙婪烙€。例如,NF-κB和STAT3通過的激活可以觸發(fā)支持體內(nèi)穩(wěn)態(tài)組織修復(fù)生長(zhǎng)因子和細(xì)胞因子的產(chǎn)生。盡管間充質(zhì)細(xì)胞是動(dòng)態(tài)的,并且對(duì)于維持體內(nèi)平衡至關(guān)重要,但是需要進(jìn)一步的工作來確認(rèn)它們是否與微生物群存在直接相互作用。
二、CRC的危險(xiǎn)因素
雖然尚未揪出CRC的真正致病元兇,但已有大量證據(jù)指出相關(guān)危險(xiǎn)因素。與炎癥性腸病(inflammatory bowel disease, IBD)相關(guān)的炎癥,以及遺傳性基因缺陷如家族性腺瘤性息肉病和Lynch綜合征是已知的遺傳危險(xiǎn)因素;數(shù)學(xué)建模將70-90%的風(fēng)險(xiǎn)歸因于環(huán)境因素,尤其是纖維含量低而紅肉含量高的飲食習(xí)慣;此外,致病性感染也是公認(rèn)的環(huán)境觸發(fā)因素,例如于1991年發(fā)現(xiàn)的幽門螺桿菌與胃癌的聯(lián)系。當(dāng)然,無論CRC的始發(fā)因素是什么,通常都涉及體內(nèi)穩(wěn)態(tài)免疫和微生物群衍生信號(hào)的破壞,刺激產(chǎn)生上皮細(xì)胞過度修復(fù)的反應(yīng)。這類反應(yīng)往往會(huì)導(dǎo)致腸道干細(xì)胞內(nèi)一連串基因突變諸如APC,KRAS或PIK3CA,以起始腫瘤發(fā)生。
腸道炎癥被認(rèn)為是CRC中的“雙刃劍”。原位T細(xì)胞的數(shù)量與CRC積極的臨床結(jié)局相關(guān),利用免疫系統(tǒng)進(jìn)行抗原特異性清除仍然是許多治療策略的目標(biāo)。然而,腫瘤通常通過免疫編輯腫瘤抗原而逃避這種免疫介導(dǎo)的破壞,使其無法被檢測(cè)到。盡管先天性免疫細(xì)胞(例如中性粒細(xì)胞和巨噬細(xì)胞)可以幫助清除腫瘤,但在某些情況下,它們會(huì)釋放活性氧,這可能通過破壞IECs的基因組完整性來引發(fā)致癌級(jí)聯(lián)反應(yīng)。因此,腫瘤細(xì)胞創(chuàng)建了一個(gè)惡性循環(huán),其中微環(huán)境中的許多細(xì)胞(包括駐留的成纖維細(xì)胞)可以被重新編程以產(chǎn)生額外的生長(zhǎng)因子,細(xì)胞因子和促血管生成因子,從而維持不受限的增殖和侵襲。
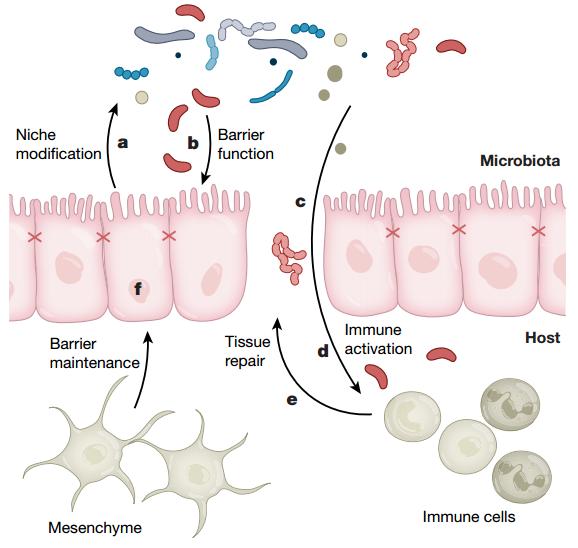
圖1,健康個(gè)體和CRC個(gè)體中宿主與腸道菌群相互作用示意圖
三、CRC患者往往伴隨腸道菌群失調(diào)
CRC中宿主與菌群的相互作用關(guān)系被初步證明于1975年,當(dāng)時(shí)觀察到致病性二甲基肼在無菌大鼠中誘發(fā)的結(jié)腸腫瘤明顯少于腸道菌群正常的大鼠。此外,相對(duì)于接受健康菌群的小鼠而言,接受CRC糞便菌群的小鼠腸息肉數(shù)量更多。由于包括16S rRNA和鳥槍法宏基因組學(xué)(shotgun metagenomics)在內(nèi)的微生物組分析技術(shù)的發(fā)展,與健康對(duì)照個(gè)體相比,CRC患者的腸道菌群成分有差異是無可厚非的,被概括為“菌群失調(diào)”。在CRC腫瘤組織中檢測(cè)出較高含量的核梭菌、大腸埃希氏菌、脆弱擬桿菌、糞腸球菌、解脲鏈球菌和消化鏈球菌菌種,而所謂的保護(hù)性菌屬包括羅氏菌、梭菌、費(fèi)卡桿菌和雙歧桿菌在內(nèi)的細(xì)菌數(shù)量明顯減少。這些差異中的一些(梭菌水平最一致)與臨床結(jié)局和化療敏感性相關(guān),因此這類細(xì)菌具有作為生物標(biāo)記物的潛力。然而,這種菌群類型和數(shù)量的變化究竟是癌癥的原因還是結(jié)果,是否能將CRC的發(fā)生歸因于某些特定的“致癌”微生物,仍具有挑戰(zhàn)性。
四、CRC相關(guān)菌群可致基因毒性
如果要定義單個(gè)微生物或群落具有致癌性,那么它們必須發(fā)揮致癌作用,比如DNA損傷。近期有許多研究報(bào)道了與CRC相關(guān)的基因毒性,其中包括產(chǎn)大腸菌素的聚酮化合物合酶(pks)+大腸桿菌(以下簡(jiǎn)寫為pks+E.coli),腸毒性脆弱性雙歧桿菌(ETBF),糞腸球菌和產(chǎn)生細(xì)胞擴(kuò)張毒素的空腸彎曲菌。尤其是pks+E.coli可致雙鏈斷裂,異倍體型和不正確的細(xì)胞分裂。在具有遺傳易感性的Il10-/-小鼠中,pks+E.coli和ETBF可以誘導(dǎo)8-氧代鳥嘌呤DNA損傷,這些損傷與結(jié)腸腫瘤的發(fā)生率具有很高的相關(guān)性。ETBF毒素可以上調(diào)精胺氧化酶水平,從而導(dǎo)致活性氧的產(chǎn)生和DNA損傷標(biāo)記γ-H2A.x的誘導(dǎo)。
五、腸道菌群驅(qū)動(dòng)的代謝物的影響
盡管飲食可以直接致癌,它同樣也可以通過改變特定物種的豐度和代謝物來影響腸道生態(tài)系統(tǒng)。例如,與高硫代謝細(xì)菌水平增加相關(guān)的飲食與遠(yuǎn)端結(jié)腸癌和直腸癌的風(fēng)險(xiǎn)增加相關(guān);高脂飲食與CRC的復(fù)發(fā)以及某些微生物的膠原蛋白水解活性相關(guān)。為鑒定與CRC相關(guān)的候選代謝物,已有研究針對(duì)Lynch綜合征和CRC患者的糞便進(jìn)行了蛋白質(zhì)組學(xué)分析。這些研究確定了氧化代謝微環(huán)境的增強(qiáng),這被認(rèn)為是由于CRC患者結(jié)腸中活性氧和活性氮的含量增加以及破壞DNA的膽汁酸脫氧膽酸的濃度增加。在缺少游離脂肪酸受體2(FFAR2)的小鼠中觀察到腫瘤發(fā)生增加,這被認(rèn)為是由于IEC完整性降低,細(xì)菌大量涌入,樹突狀細(xì)胞過度活化以及CD8 + T細(xì)胞表型耗竭所致。
六、炎癥反應(yīng)在CRC中的作用
炎癥是結(jié)直腸癌發(fā)生的公認(rèn)驅(qū)動(dòng)因素,因此患有IBD的個(gè)體患CRC的風(fēng)險(xiǎn)增加。然而,菌群塑造腫瘤免疫環(huán)境的途徑,進(jìn)而改變周圍微生物群的方式,是正在進(jìn)行的研究的重點(diǎn)。腸道上皮屏障的破壞使得被分隔的微生物流入組織。有研究報(bào)道,缺乏模式識(shí)別受體相關(guān)基因Nod1,Nod2和Ripk2的小鼠腫瘤多樣性和IEC通透性增強(qiáng),強(qiáng)調(diào)宿主防御和IEC功能在預(yù)防腫瘤發(fā)生中的重要性。
由腸道屏障缺陷導(dǎo)致的細(xì)菌轉(zhuǎn)移與多種癌癥相關(guān)促炎因子的產(chǎn)生相關(guān),包括IL-1β,IL-23,IL-22,IL-27,尤其是IL-17A和IL-6。與此相一致,在小鼠和人類中,發(fā)現(xiàn)有細(xì)菌浸潤(rùn)的腫瘤中,IL-6,IL-23和IL-17A的表達(dá)均發(fā)生上調(diào)。此外,IL-6與IEC上的受體結(jié)合以觸發(fā)異常增殖。此外,受體表達(dá)模式的重要性通過一項(xiàng)研究得到例證,該研究表明,IL-1受體的CD4+ T細(xì)胞特異性消融減少了腫瘤誘發(fā)的炎癥反應(yīng),而髓樣細(xì)胞該受體的缺乏則會(huì)增加細(xì)菌入侵和IL-17A分泌,以及增殖腫瘤的數(shù)量和大小。
值得注意的是,許多細(xì)胞因子在CRC中顯示多效性功能。例如,已知由菌群觸發(fā)并由T細(xì)胞和固有淋巴樣細(xì)胞產(chǎn)生的IL-22可以維持IEC增殖并在炎癥過程中誘導(dǎo)抗凋亡蛋白的表達(dá)(促腫瘤發(fā)生的作用),但也可以在基因毒性損傷后,誘導(dǎo)屏障完整性和DNA損傷修復(fù)(抗腫瘤發(fā)生的作用)。當(dāng)然,這些機(jī)制中的部分結(jié)論是從使用小鼠適應(yīng)性病原體(例如肝螺桿菌)的模型系統(tǒng)中獲得的。細(xì)胞因子在致癌過程中的不同作用取決于它們?cè)诩膊∵M(jìn)展過程中的濃度和時(shí)間動(dòng)態(tài)性,以及其他細(xì)胞類型,細(xì)胞因子等背景,而這些變量經(jīng)常在許多還原研究中被忽略。
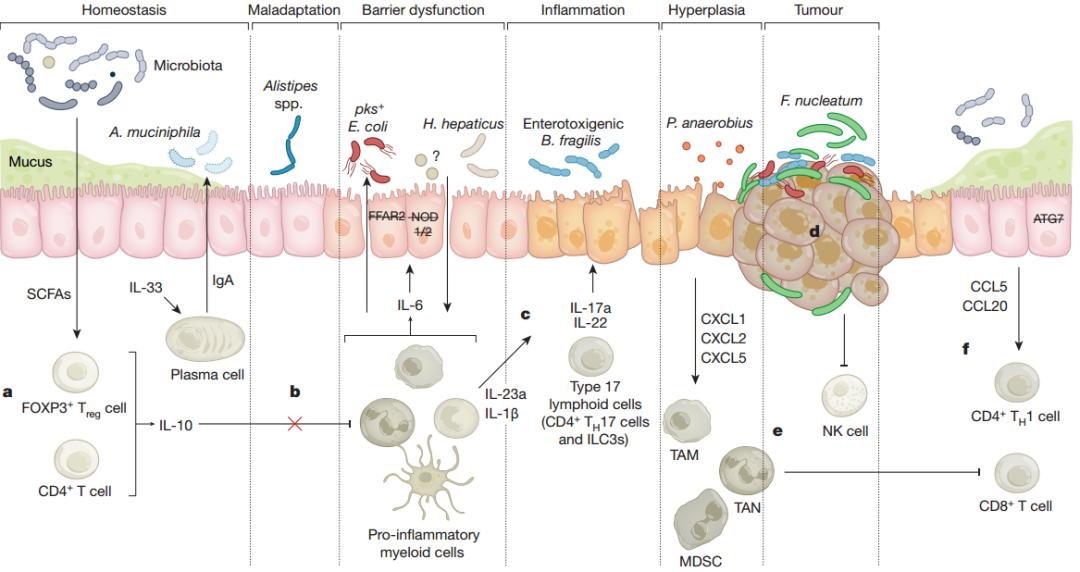
圖2,腸道菌群促成CRC的已知炎癥機(jī)制
七、炎癥驅(qū)動(dòng)的微生物生態(tài)位
對(duì)IBD的研究已經(jīng)說明,不受控制的炎癥會(huì)在結(jié)腸內(nèi)產(chǎn)生不同的生態(tài)位(ecological niche),從而導(dǎo)致更適應(yīng)的菌群存活。盡管這通常被認(rèn)為是有害的,但這種細(xì)微差別的過程可能在進(jìn)化上很重要。
特定“致癌”物種的豐富程度似乎部分取決于抗炎細(xì)胞因子IL-10,這說明了致癌過程中免疫適應(yīng)不良的有害后果。的確,在IL-10敲除小鼠的腸中觀察到基因毒性的pks+E.coli數(shù)量增加,且IL-10缺失與擬桿菌屬,普氏桿菌屬和里克氏菌屬增加的豐度相一致。重要的是,菌群可以例如通過產(chǎn)生丁酸鹽以反饋從而改變IL-10的水平。丁酸酯特異性受體GPR109A的下游信號(hào)可以抑制炎癥驅(qū)動(dòng)的致癌作用,并使巨噬細(xì)胞和樹突狀細(xì)胞使CD4+ T細(xì)胞偏向產(chǎn)生IL-10的T調(diào)節(jié)細(xì)胞。類似于IL-10,IL-33的缺失也會(huì)增強(qiáng)對(duì)炎癥驅(qū)動(dòng)的結(jié)腸癌的敏感性。值得注意的是,將IL-18缺陷型和野生型小鼠共居以實(shí)現(xiàn)菌群轉(zhuǎn)移,這一操作引起野生型小鼠中炎癥誘導(dǎo)的CRC患病率增加。相反,一些研究報(bào)道IL-18可能通過抑制產(chǎn)粘液的杯狀細(xì)胞成熟來促進(jìn)DSS誘導(dǎo)的結(jié)腸炎。這些看似矛盾的數(shù)據(jù)也可能是由于不同的時(shí)空因素,劑量因素等造成,但關(guān)于這點(diǎn)還未能得到明確結(jié)論。
八、“致癌微生物”引起免疫成分改變
腫瘤可以通過募集特定的免疫細(xì)胞來創(chuàng)建適宜腫瘤生長(zhǎng)的微環(huán)境,其密度和成分能被菌群所影響。例如,吸引淋巴細(xì)胞的趨化因子CCL5、CCL20和CXCL11的水平與擬桿菌和厚壁菌門的成員相關(guān),并且可以在體外被具核梭桿菌和大腸桿菌誘導(dǎo)。然而,這些化學(xué)誘導(dǎo)物是否募集保護(hù)性或致病性T細(xì)胞亞群尚不清楚,可能會(huì)受到基因背景的影響,在IECs中缺乏自噬調(diào)節(jié)基因Atg7的小鼠中,菌群與細(xì)胞毒性CD8+ T細(xì)胞和CD4+ TH1反應(yīng)相關(guān)。因此,改變菌群以使其傾向于“熱”腫瘤微環(huán)境(通常以浸潤(rùn)C(jī)D8+ T細(xì)胞和患者有益的結(jié)果為特征)將是一種有趣的治療方案。
ETBF已被發(fā)現(xiàn)會(huì)在ApcMin /+小鼠中由STAT3激活驅(qū)動(dòng)誘發(fā)促癌TH17的反應(yīng)。這伴隨著iNoshigh單核細(xì)胞樣髓樣來源的抑制細(xì)胞(M-MDSCs)的募集和分化,腫瘤內(nèi)IL-17A與IL-17R +髓樣細(xì)胞結(jié)合驅(qū)動(dòng)Nos2上調(diào)。這些M-MDSC抑制了細(xì)胞毒性CD8 + T細(xì)胞的活性,同時(shí)誘導(dǎo)了涉及腫瘤生長(zhǎng)(Mmp9)和血管生成(Vegfa)的基因表達(dá)。
與ETBF相似,在DSS誘導(dǎo)的CRC小鼠模型中用CRC相關(guān)溶脂鏈球菌進(jìn)行管飼會(huì)導(dǎo)致腫瘤負(fù)荷增加,免疫抑制性CD11b +髓樣細(xì)胞的選擇性募集和髓樣來源的細(xì)胞因子(包括IL-6和IL-8)水平升高。喂食具核梭桿菌的ApcMin / +小鼠的腫瘤負(fù)荷更大,同時(shí)伴隨著免疫抑制性瘤內(nèi)髓樣細(xì)胞的增多。據(jù)報(bào)道,與CRC相關(guān)的厭氧消化鏈球菌可通過選擇性粘附于結(jié)腸癌細(xì)胞并誘導(dǎo)NF-κb來觸發(fā)ApcMin / +小鼠中促腫瘤髓細(xì)胞群體的擴(kuò)張,以及T調(diào)節(jié)細(xì)胞,TH17和細(xì)胞毒性CD8 + T細(xì)胞頻率同時(shí)增加。具核梭桿菌還可以以不依賴細(xì)胞因子的方式起作用,并直接抑制自然殺傷細(xì)胞的細(xì)胞毒性,從而使腫瘤能夠逃避免疫系統(tǒng)追殺。用具核梭桿菌預(yù)處理CRC細(xì)胞系通過受體TIGIT與具核梭桿菌衍生蛋白Fap2的結(jié)合降低了共培養(yǎng)的NK細(xì)胞的活性。
九、研究“致癌微生物”的新興技術(shù)
綜上所述,盡管在這一領(lǐng)域已經(jīng)取得了實(shí)質(zhì)性的進(jìn)展,但尚且缺乏關(guān)于改變的菌群與宿主之間在治療上存在因果作用的證據(jù)。編撰一部完整的與CRC相關(guān)的包括細(xì)菌、真菌、病毒、古細(xì)菌和代謝物的豐度和功能菌群譜,并考慮這些因素與宿主之間的多層串?dāng)_是一項(xiàng)尚待解決的艱巨任務(wù),而這部分是由于測(cè)序深度的限制,特定菌種難以培養(yǎng)以及復(fù)現(xiàn)人類腫瘤微環(huán)境模型的局限性等。
首先,類器官是上皮組織的三維培養(yǎng)系統(tǒng),可在體外穩(wěn)定培養(yǎng)源自患者的腫瘤組織。通過使用該技術(shù),可以測(cè)試各種刺激效果,但是對(duì)類器官封閉頂端表面的接近性較差,使得具有菌群成分的培養(yǎng)具有挑戰(zhàn)性。緊接著,細(xì)菌的類器官內(nèi)顯微注射技術(shù)的發(fā)展和上皮極性逆轉(zhuǎn)的研究已經(jīng)開始解決這個(gè)問題。例如,將pks+E.coli注射至類器官中揭示了這種細(xì)菌直接引起臨床相關(guān)的CRC驅(qū)動(dòng)突變的能力。盡管這種還原論方法有自己的價(jià)值,但重要的是要了解致病菌群對(duì)基因背景以及來自腫瘤微環(huán)境的其他細(xì)胞起作用的能力。因此,科研人員將類器官實(shí)驗(yàn)擴(kuò)展到與免疫細(xì)胞和間充質(zhì)細(xì)胞共培養(yǎng),使用CRISPR–Cas9引入靶向突變。
進(jìn)一步的,科研人員開發(fā)了一種稱為“器官芯片(organ-on-a-chip)”的體外微流系統(tǒng),該系統(tǒng)利用特定的通道滲透性整合了多個(gè)細(xì)胞元素。這種微流體模型稱為“HuMIX”,通過共培養(yǎng)營(yíng)養(yǎng)物質(zhì),活微生物和人上皮細(xì)胞來產(chǎn)生獨(dú)特的短鏈脂肪酸(short-chain fatty acids, SCFAs)比率,限制IECs的自我更新能力,從而提供了與CRC相關(guān)代謝產(chǎn)物的功能性見解。重要的是,該技術(shù)已經(jīng)擴(kuò)展到可以對(duì)厭氧和好氧菌群進(jìn)行建模,這些進(jìn)展還將有助于研究微生物內(nèi)部對(duì)必需營(yíng)養(yǎng)素的競(jìng)爭(zhēng),可能與致癌微生物的生長(zhǎng)相關(guān)。
基因工程小鼠模型(Genetically engineered mouse models, GEMM)構(gòu)成了這篇綜述中討論的許多研究的基礎(chǔ)。然而,一些模型潛伏期較長(zhǎng)且不能復(fù)現(xiàn)人類疾病的晚期階段。為了減少模型的持續(xù)時(shí)間,從而在不發(fā)生細(xì)菌抗藥性的情況下進(jìn)行抗生素處理,可以將致瘤類器官局部注入實(shí)驗(yàn)動(dòng)物的直腸。對(duì)于在體內(nèi)還原微生物-宿主相互作用的復(fù)雜性以及證明CRC的致病因果關(guān)系。為了研究晚期CRC,已經(jīng)設(shè)計(jì)出轉(zhuǎn)移模型,并提供了工具來研究菌群在調(diào)節(jié)上皮-間質(zhì)轉(zhuǎn)化中的潛在作用。
值得注意的是,盡管抗生素是在疾病發(fā)展過程中調(diào)節(jié)微生物群的有用工具,但它們通常針對(duì)某些微生物而沒有足夠的控制或確保根除,從而使數(shù)據(jù)解釋具有挑戰(zhàn)性。例如,在DSS-乙氧基甲烷癌癥模型中,在無菌小鼠中觀察到增強(qiáng)的腫瘤發(fā)生,有趣的是,使用抗生素處理的結(jié)果卻相反。這種看似相反的數(shù)據(jù)可能是由于具有抗生素抗性菌群的存在,或消除菌群的時(shí)間點(diǎn)不同再或者無菌小鼠的免疫系統(tǒng)不同等因素。除細(xì)菌和代謝產(chǎn)物外,有越來越多的證據(jù)表明真菌,古細(xì)菌和病毒也在CRC中發(fā)生改變。最近有通過真菌共生體誘導(dǎo)IL-18可抑制結(jié)腸炎相關(guān)的小鼠CRC,從而突顯其保護(hù)性作用。
對(duì)于這樣的集成分析流水線,廣泛而系統(tǒng)的“多組學(xué)(multi-omic)”方法對(duì)于構(gòu)建交互組(interactome)至關(guān)重要,另一個(gè)新興領(lǐng)域是在菌群背景下對(duì)表觀遺傳學(xué)進(jìn)行分析,例如與CRC腫瘤中獨(dú)特的甲基化模式相關(guān)的梭桿菌屬。當(dāng)然,鑒于模型系統(tǒng)之間存在廣泛的個(gè)體異質(zhì)性和局限性,因此大規(guī)模國(guó)際患者隊(duì)列(包括有關(guān)宿主的菌群和遺傳特征的綜合數(shù)據(jù))對(duì)于確定臨床相關(guān)發(fā)現(xiàn)至關(guān)重要。
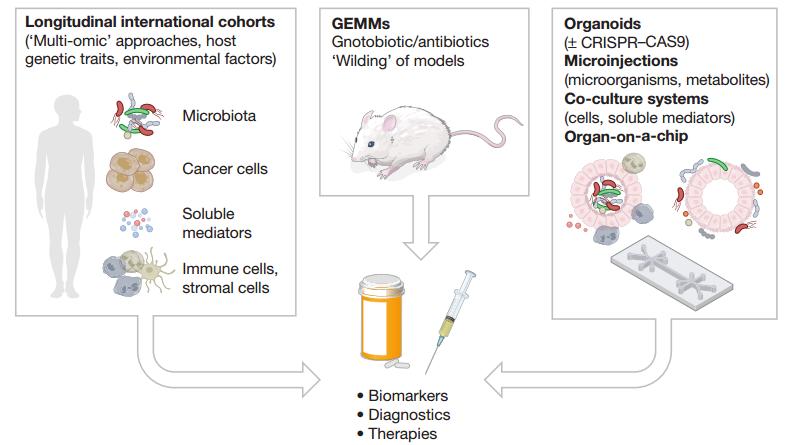
圖3,研究腸道菌群在CRC中致病作用的方法學(xué)
討論
在過去的十年中迅速累積的研究發(fā)現(xiàn),早期癌前病變的個(gè)體與轉(zhuǎn)移性CRC個(gè)體的腸道菌群組成存在差異。在CRC中鑒定出的具核梭桿菌,大腸桿菌或脆弱擬桿菌的豐度增加,開創(chuàng)了腫瘤研究的新領(lǐng)域,即微生物可以作為生物標(biāo)記物與常規(guī)診斷方法結(jié)合使用。靶向菌群的治療方法也正在興起,包括選擇性消除致癌菌群,糞便移植抗致癌菌群以及通過口服補(bǔ)充劑或靶向噬菌體療法使微生物群傾斜。然而,由于微生物組對(duì)宿主生物學(xué)的多種作用,因此需要謹(jǐn)慎考慮上述治療手段是否存在副作用。
非常重要的是,當(dāng)前必須破譯某一特定菌株或“致癌”菌群(包括細(xì)菌,真菌,病毒,古細(xì)菌及其產(chǎn)生的代謝產(chǎn)物)的聯(lián)合是否會(huì)改變疾病狀態(tài)。由于微生物群中個(gè)體內(nèi)部的異質(zhì)性很大,那么可以認(rèn)為,不同個(gè)體中的不同菌株將通過共同途徑以觸發(fā)相似的病理改變,因此,針對(duì)廣泛而非特定的單元研究可能是最有效的策略。
總而言之,這篇綜述已經(jīng)向我們展示了截至目前針對(duì)腸道菌群對(duì)宿主CRC的各種可能機(jī)制,從改變腸道屏障和/或免疫環(huán)境到重塑結(jié)腸生態(tài)位和提供基因毒性損傷等。盡管宿主的適應(yīng)不良和屏障破壞對(duì)于促進(jìn)宿主與微生物群之間的緊密接觸非常關(guān)鍵,但仍需要縱向多組學(xué)來解釋這些“階段”在腫瘤發(fā)展中的時(shí)間順序。當(dāng)然,非常幸運(yùn)的是,目前已有大批新興技術(shù)為在模擬生理CRC系統(tǒng)的背景下研究微生物群,這對(duì)于破解結(jié)腸腫瘤微環(huán)境的挑戰(zhàn)性和復(fù)雜性提供可能。
原文鏈接:
https://doi.org/10.1038/s41586-020-2729-3
來源:BioArt



1980-2020
原標(biāo)題:《【學(xué)術(shù)前沿】Nature綜述 | 腸道菌群與結(jié)直腸癌的研究進(jìn)展》
閱讀原文
 魯ICP備2020050029號(hào)-1
魯ICP備2020050029號(hào)-1 魯ICP備2020050029號(hào)-1
魯ICP備2020050029號(hào)-1